- 弱化现象

弱化现象
化学弱化
弱化
弱化,即变弱;使变弱。[2]
定义
英文名称:attenuation
定义1:由于基因内部弱化子的作用,提前终止转录而抑制基因表达。是细菌控制操纵子表达的转录调节机制之一,见于合成氨基酸等生物小分子的操纵子。
应用学科:生物化学与分子生物学(一级学科);基因表达与调控(二级学科)
定义2:降低细菌操纵子转录效率并提前终止转录的一种调控机制。
应用学科:遗传学(一级学科);分子遗传学(二级学科)
操纵子
名词定义
操纵子(operon)指包含结构基因、操纵基因以及启动基因的一些相邻基因组成的DNA片段,其中结构基因的表达受到操纵基因的调控。主要见于原核生物,但在真核生物中也存在。
基本结构
大肠杆菌色氨酸操纵子结构较简单,也是研究得最清楚的操纵子,结构基因依次排列为trpE,trpD,trpC,trpB,trpA,其中trpG与trpD 和trpC与trpF分别发生基因融合。trpE和trpG编码邻氨基苯甲酸合酶,trpD编码邻氨基苯甲酸磷酸核糖转移酶,trpC编码吲哚甘油磷酸合酶,trpF编码异构酶,trpA和trpB分别编码色氨酸合酶的α和β亚基。trpE的上游为调控区,由启动子、操纵基因和162bp 的前导序列组成。5 个结构基因全长约6800bp,trpD远侧还有一个二级启动子,在细胞生长需要过量Trp时发挥作用。
一些G+菌,如枯草杆菌色氨酸操纵子的结构有所不同,7 个结构基因中的6 个依次排列为trpE,trpD,trpC,trpF,trpB,trpA,存在于含有12个结构基因的芳香族氨基酸超操纵子( aro operon ),第7 个结构基因,trpG存在于叶酸合成操纵子中,该酶参与Trp 和叶酸的合成。有2个启动子参与调控,一个位于aro operon的起始位置,另一个则位于trpE 上游约200 bp处。
调控作用
Trp合成途径较漫长,消耗大量能量和前体物,如丝氨酸、PRPP、谷氨酰氨等,是细胞内最昂贵的代谢途径之一,因此受到严格调控,其中色氨酸操纵子发挥着关键作用。调控作用主要有三种方式:阻遏作用、弱化作用以及终产物Trp 对合成酶的反馈抑制作用。
阻遏作用
trp操纵子转录起始的调控是通过阻遏蛋白实现的。产生阻遏蛋白的基因是trpR,该基因距trp operon基因簇很远。它结合于trp 操纵基因特异序列,阻止转录起始。但阻遏蛋白的DNA结合活性受Trp调控,Trp起着一个效应分子的作用,Trp与之结合的动力学常数为1~2 ×10- 5mol·L - 1。在有高浓度Trp存在时,阻遏蛋白- 色氨酸复合物形成一个同源二聚体,并且与色氨酸操纵子紧密结合,因此可以阻止转录。阻遏蛋白- 色氨酸复合物与基因特异位点结合的能力很强,动力学常数为2 ×10- 10mol·L - 1,因此细胞内阻遏蛋白数量仅有20~30分子已可充分发挥作用。当Trp 水平低时,阻遏蛋白以一种非活性形式存在,不能结合DNA。在这样的条件下,trp操纵子被RNA聚合酶转录,同时Trp 生物合成途径被激活。
弱化作用
trp操纵子转录终止的调控是通过弱化作用( attenuation)实现的。在大肠杆菌trp operon,前导区的碱基序列包括4个分别以1、2、3和4表示的片段,能以两种不同的方式进行碱基配对,1 - 2和3 -4配对,或2 - 3配对,3 - 4配对区正好位于终止密码子的识别区。前导序列有相邻的两个色氨酸密码子,当培养基中Trp 浓度很低时,负载有Trp 的tRNATrp也就少,这样翻译通过两个相邻色氨酸密码子的速度就会很慢,当4区被转录完成时,核糖体滞留1区,这时的前导区结构是2 - 3配对,不形成3 - 4配对的终止结构,所以转录可继续进行。反之,核糖体可顺利通过两个相邻的色氨酸密码子,在4区被转录之前,核糖体就到达2区,这样使2 - 3不能配对,3 - 4 区可以配对形成终止子结构,转录停止。
枯草杆菌的弱化作用机制另有特点。因其色氨酸操纵子结构的特殊性,转录起始的调节似乎不如转录终止的调节更具重要性。枯草杆菌色氨酸操纵子表达主要受到色氨酸激活RNA结合蛋白( Trp -activated RNA - binding p rotein,TRAP)的调节。该蛋白与色氨酸结合被激活后,可与trpE上游转录产物结合,导致转录终止。当色氨酸浓度较低时,TRAP失活,转录可以继续,结构基因得以表达。另外枯草杆菌对未负荷色氨酸的tRNATrp也很敏感,后者大量堆积,会诱导合成抗TRAP 蛋白( anti -TRAP,AT)。AT与Trp激活的TRAP结合,可以取消其转录终止活性。trpG表达也受TRAP调控,活化的TRAP与和trpG相重叠的S - D 序列结合,阻碍核糖体的结合,抑制trpG转录。
反馈抑制作用
由于基因表达必然消耗一定的能源和前体物,相对于阻遏和弱化作用,反馈抑制作用更为经济和高效。终产物Trp对催化分支途径几步反应的酶具有反馈抑制作用,其50%抑制浓度分别为:邻氨基苯甲酸合酶,0. 0015 mmol·L - 1 ;邻氨基苯甲酸磷酸核糖转移酶,0. 15 mmol·L - 1 ; 色氨酸合成酶,7. 7mmol·L - 1。对于普通野生菌株,邻氨基苯甲酸合酶对Trp合成起到关键调控作用,常被称为瓶颈酶;但对高产Trp工程菌而言,上述任何一种酶的反馈抑制都会直接影响Trp产量。研究发现酶蛋白某些特殊位点突变可以导致对反馈抑制作用敏感性显著下降,如邻氨基苯甲酸合酶38位的丝氨酸被精氨酸取代,抗反馈抑制能力显著提高,当环境中Trp浓度为10 mmol·L - 1时酶活性不受影响,而相同条件下野生型酶活性不到1%。邻氨基苯甲酸磷酸核糖转移酶162位缬氨酸被谷氨酸取代,抗反馈抑制能力也有显著提高,当环境中含有0. 83 mmol·L - 1色氨酸或0. 32 mmol·L - 1 5 - 甲基- 色氨酸时,酶活性分别为野生菌的3. 6倍和2. 4倍。陈小芳等报道一株谷氨酸棒杆菌邻氨基苯甲酸合酶基因7个碱基突变导致6个氨基酸残基改变,抗反馈抑制能力显著增强,环境中Trp 浓度达到15 mmol·L - 1时,邻氨基苯甲酸合酶活性几乎没有变化。
遗传改造
由于色氨酸操纵子的调控作用,自然界不可能存在高产Trp菌株,为了获得高产Trp菌株,就必须对色氨酸操纵子进行改造,解除其调节作用。早期的研究策略主要依靠传统诱变方法,经过长期努力,获得了一些有价值的研究结果,如获得了TrpR - 菌株,通过缺失某些片断解除了弱化作用,得到了一些抗反馈抑制的酶。许多Trp生产菌株都是通过随机的诱变技术筛选得到的,如王健等通过硫酸二乙酯诱变,Trp 类似物筛选等方法从谷氨酸棒杆菌中培育出一株trp 高产菌株,摇瓶发酵64 h,产trp达到7. 28 g·L - 1。
传统诱变的方法尽管有效,但其缺陷点也是显而易见的,如工作量大,效率低,突变株的菌体生长、对环境的耐受性以及遗传稳定性等都比野生型菌株差等。基因工程技术的建立和发展对色氨酸操纵子改造提供了新的技术平台。1979年Tribe等人采用DNA重组技术对大肠杆菌进行改造,扩增trp 操纵子,发酵12 h,产酸1 g·L - 1,产酸量尽管不是很高,但是其意义却十分重大,由此开创了基因工程技术在Trp生物合成应用的先河。随后,Aiba等将带有色氨酸操纵子的质粒引入大肠杆菌,发酵27 h,并补充邻氨基苯甲酸,得到trp 6. 2 g·L - 1。Ikeda等通过构建稳定质粒,扩增分支途径限速酶并改造中心代谢途径,获得产Trp 达58 mg ·L - 1 的菌株。除了扩增表达操纵子基因,对其进行理性设计和改造也开始引起关注。已知酶分子某些特殊位点突变可以导致对反馈抑制作用敏感性下降,因此可以考虑利用基因工程技术对色氨酸操纵子结构基因进行理性改造降低其对反馈抑制的敏感性,但是Z至今尚缺乏成功的范例,主要原因在于现有酶分子反馈抑制结构与功能关系资料不足,不能满足需要。
理论研究
1991年,Bailey用代谢工程描述利用DNA重组技术对细胞的酶反应、物质运输以及调控功能的遗传操作,进而改良细胞生物活性的过程,标志着代谢工程向一门系统学科发展的转折点。代谢工程亦称途径工程,以区别于传统的单基因表达(第一代基因工程)和基因定向突变(第二代基因工程),是有目的地对细胞生化反应的代谢网络进行修饰的技术,在多基因水平上设计修饰细胞固有的代谢途径和遗传性状,并赋予细胞更为优越甚至崭新的产物生产品质。代谢工程在提高宿主细胞原有代谢物的产量、产生新物质、扩展和构建新代谢途径、生产代谢产物如氨基酸、抗生素、维生素以及降解环境污染物等诸多方面显示出广阔的应用前景。从理论上提高Trp产率是代谢工程的首要任务,这需要对Trp生物合成和对细胞内控制Trp 代谢的异化途径有很好的了解,同时还要有一个在较宽的微生物代谢网络内描述这些途径的有效的数学模型。早期的模型主要考虑色氨酸操纵子动力学的某个方面,仅有少数研究模型,综合考虑了色氨酸操纵子的三种作用机制。修志龙等将代谢工程理论引入trp代谢分析领域,建立了适宜的数学模型,发现在代谢稳定的条件下,阻遏水平和酶的反馈抑制强度严重地影响了目标变量,即trp 浓度。Santillan等人提出的动力模型采用Second Lyapunov’s method分析,通过对野生菌株和几株改良菌株(邻氨基苯甲酸合成酶反馈抑制和弱化作用分别解除)的性能进行比较、验证,得出结论认为酶的反馈抑制对于系统稳定性具有重要作用,而弱化作用影响较小,主要在Trp营养发生改变时发生作用。这两个模型有一定的代表性,它们考虑了酶的反馈抑制,对于Trp生物合成具有一定指导意义;但其不足也很明显,仅仅考虑了邻氨基苯甲酸合成酶的反馈抑制作用,对其它酶未作考虑,另外一个不足是缺乏高产色氨酸菌株来加以验证。
操纵子学说
1961年,法国科学家莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)发表“蛋白质合成中的遗传调节机制”一文,提出操纵子学说,开创了基因调控的研究。
学说介绍
1961年,法国科学家莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)发表“蛋白质合成中的遗传调节机制”一文,提出操纵子学说,开创了基因调控的研究。四年后的1965年,莫诺与雅可布即荣获诺贝尔生理学与医学奖。
莫诺与雅可布最初发现的是大肠杆菌的乳糖操纵子。这是一个十分巧妙的自动控制系统,这个自动控制系统负责调控大肠杆菌的乳糖代谢。
乳糖可作为培养大肠杆菌的能源。大肠杆菌能产生一种酶(叫做“半乳糖苷酶”),能够催化乳糖分解为半乳糖和葡萄糖,以便作进一步的代谢利用。编码半乳糖苷酶的基因(简称z)是一个结构基因(structural gene)。这个结构基因与操纵基因共同组成操纵子。操纵基因受一种叫作阻遏蛋白的蛋白质的调控。当阻遏蛋白结合到操纵基因之上时,乳糖会起诱导作用,它与阻遏蛋白结合,使之从操纵基因上脱落下来。这时,操纵基因开启,相邻的结构基因也表现活性,细菌就能分解并利用乳糖了,这样,乳糖便成了诱导半乳糖苷酶产生的诱导物。
上述内容表明,大肠杆菌的乳糖操纵子是一个十分巧妙的自动控制系统:当培养基中含有充分的乳糖,同时不含葡萄糖时,细菌便会自动产生半乳糖苷酶来分解乳糖,以资利用。当培养基中不含乳糖时,细菌便自动关闭乳糖操纵子,以免浪费物质和能量。
启动基因
60年代中期,在操纵子中还发现了另一个开关基因,称为启动基因(promoter)。启动基因位于操纵基因之前,二者紧密相邻。启动基因由环腺苷酸(cAMP)启动,而环腺苷酸能被葡萄糖所抑制。这样,葡萄糖便通过抑制环腺苷酸而间接抑制启动基因,使结构基因失活,停止合成半乳糖苷酶。
由此可知,结构基因同时受两个开关基因——操纵基因与启动基因的调控。只有当这两个开关都处于开启状态时,结构基因才能活化。当培养基中同时存在葡萄糖和乳糖时,葡萄糖通过抑制环腺苷酸而间接抑制启动基因,并进而抑制结构基因,使细菌不产生半乳糖苷酶。这种情况下,细菌便会自动优先利用葡萄糖,因为葡萄糖果是比乳糖更好的能源。
1969年,贝克维斯(J·R·Beckwith)从大肠杆菌的DNA中分离出乳糖操纵子,完全证实了雅可布和莫诺的模型。
在启动基因发现之前,莫诺和雅可布的操纵子模型中,直接对结构基因起操纵作用的开关基因,仅有一个操纵基因。因此,有人开玩笑说:“半个操纵子就可以得诺贝尔奖”。对某一项成就,人们如果说它的一半就可以实现某种重要作用,就表明这项成就的伟大。我国北宋时代的名臣赵普就有“半部论语就可以治天下”的名言,由此也可见操纵子学说的巨大意义。
操纵子模型
一个控制细胞基因表达的模型称为操纵子(operon),此模型的提出使基因概念又向前迈出了一大步。表明人们已认识到基因的功能并不是固定不变的,而是可以根据环境的变化进行调节。随之人们发现无论是真核还是原核生物转录调节都是涉及到编码蛋白的基因和DNA上的元件。这一发现获得了1965年诺贝尔奖。一个基因就是一段编码有功能产物的DNA顺序。基因的产物可以是蛋白质或是RNA(如tRNA和rRNA)。基因的重要特点是在有的情况下其产物能从合成位点散开去作用别的位点。
DNA元件是DNA上一段顺序,它不能转变成任何其它的形式,但它作为一种原位(in situ)顺序具有特殊的功能。由于它只能作用同一条DNA,因此称顺式作用元件(cis-acting element)。
基因可以根据它们的产物分成不同的类型。编码细胞必要的蛋白,如酶或结构蛋白的基因称为结构基因( structural genes)。这类基因在细胞中占绝大部分,承担着细胞各种蛋白的结构和功能。编码调节蛋白的基因称调节基因(regulator genes)。调节蛋白可调节其它基因的表达。由于调节基因的产物可以自由地结合到其相应的靶上,因此被为反式作用因子(trans-acting因子)。
调节的关键是调节基因编码调节蛋白,此蛋白通过和DNA上特殊位点的结合来控制转录。顺式作用元件被反式作用因子识别可以以“正”的或“负”的形式调节靶基因,顺式作用位点通常总是在靶基因的上游。
弱化现象
trp操纵子
trp操纵子是负责色氨酸合成的操纵子。trp操纵子是由一个启动子和一个操纵基因区组成。该操纵基因控制一个编码色氨酸生物合成需要的5种蛋白的多顺反子mRNA的表达。
发现现象
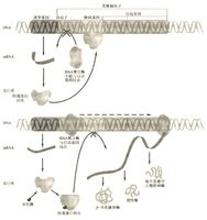 弱化现象(5)弱化现象(attenuation)是指大肠杆菌的色氨酸操纵子表达中发现的现象,在trp mRNA 5,端trp正基因的起始密码前有一个长162 bp的DNA序列称为前导区,其中第123~150位核苷酸如果缺失,trp基因的表达水平可提高6倍。研究发现,当mRNA开始合成后,除非培养基中完全不含色氨酸,否则转录总是在这个区域终止,产生一个仅有140个核苷酸的RNA分子,终止trp基因转录。换句话说,当123~150位序列缺失后,trp基因转录就不会中途终止,于是打户基因表达水平提高。123~150位序列终止转录的作用是可以被调控的,如在培养基中完全不含色氨酸,则转录不会终止,这个区域被称为弱化子。弱化现象在原核生物中是相当普遍的,大肠杆菌和鼠伤寒沙门氏菌中已陆续发现不少操纵子都有弱化现象。
弱化现象(5)弱化现象(attenuation)是指大肠杆菌的色氨酸操纵子表达中发现的现象,在trp mRNA 5,端trp正基因的起始密码前有一个长162 bp的DNA序列称为前导区,其中第123~150位核苷酸如果缺失,trp基因的表达水平可提高6倍。研究发现,当mRNA开始合成后,除非培养基中完全不含色氨酸,否则转录总是在这个区域终止,产生一个仅有140个核苷酸的RNA分子,终止trp基因转录。换句话说,当123~150位序列缺失后,trp基因转录就不会中途终止,于是打户基因表达水平提高。123~150位序列终止转录的作用是可以被调控的,如在培养基中完全不含色氨酸,则转录不会终止,这个区域被称为弱化子。弱化现象在原核生物中是相当普遍的,大肠杆菌和鼠伤寒沙门氏菌中已陆续发现不少操纵子都有弱化现象。
分析前导肽序列,发现它包括起始密码子AUG和终止密码子UGA,编码了一个14个氨基酸的多肽。该多肽有一个特征,其第10位和11位有相邻的两个色氨酸密码子。正是这两个相连的色氨酸密码子(组氨酸、苯丙氨酸操纵子中都有这种现象)调控了蛋白质的合成。
当培养基中色氨酸的浓度很低时,负载有色氨酸的tRNATrp也就少,这样翻译通过两个相邻色氨酸密码子的速度就会很慢,当4区被转录完成时,核糖体才进行到1区(或停留在两个相邻的trp密码子处),这时的前导区结构是2-3配对,不形成3-4配对的终止结构,所以转录可继续进行,直到将trp操纵子中的结构基因全部转录。
当培养基中色氨酸浓度较高时,核糖体可顺利通过两个相邻的色氨酸密码子,在4区被转录之前就到达2区,使2-3区不能配对,3-4区自由配对形成基一环终止子结构,转录被终止,trp操纵子被关闭。
合成步骤
色氨酸分5步完成。每个环节需要一种酶,编码这5种酶的基因紧密连锁在一起,被转录在一条多顺反子mRNA上,分别以trpE、trpD、trpC、trpB、trpA代表,编码了邻氨基苯甲酸合成酶、邻氨基苯甲酸焦磷酸转移酶、邻氨基苯甲酸异构酶、色氨酸合成酶和吲哚甘油-3-磷酶合成酶。
trpE基因是第一个被翻译的基因,和trpL和trpa(不是trpA)。trp操纵子中产生阻遏物的基因是trpR,该基因距trp基因簇很远,后者在大肠杆菌染色体图上25min处,而前者则位于90min处。在位于65min处还有一个trpS(色氨酸tRNA合成酶),它和携带有trp的tRNATrp也参与trp操纵子的调控作用。
基本结构
大肠杆菌色氨酸操纵子结构较简单,也是研究得最清楚的操纵子,结构基因依次排列为trpEDC2BA ,其中trpGD 和trpCF 基因融合。trpE和trpG编码邻氨基苯甲酸合酶, trpD编码邻氨基苯甲酸磷酸核糖转移酶, trpC编码吲哚甘油磷酸合酶, trpF编码异构酶, trpA和trpB分别编码色氨酸合酶的α和β亚基。trpE的上游为调控区,由启动子、操纵基因和162bp 的前导序列组成。5 个结构基因全长约6800bp, trpD远侧还有一个二级启动子,在细胞生长需要过量Trp时发挥作用。一些G 菌,如枯草杆菌色氨酸操纵子的结构有所不同, 7 个结构基因中的6 个依次排列为trpEDCFBA ,存在于含有12个结构基因的芳香族氨基酸超操纵子( a ro operon ) ,第7 个结构基因, trpG存在于叶酸合成操纵子中,该酶参与Trp 和叶酸的合成。有2个启动子参与调控,一个位于aro operon的起始位置, 另一个则位于trpE 上游约200 bp处。
调控途径
Trp合成途径较漫长,消耗大量能量和前体物,如丝氨酸、PRPP、谷氨酰氨等,是细胞内最昂贵的代谢途径之一,因此受到严格调控,其中色氨酸操纵子发挥着关键作用。调控作用主要有三种方式:阻遏作用、弱化作用以及终产物Trp 对合成酶的反馈抑制作用。
阻遏作用
trp操纵子转录起始的调控是通过阻遏蛋白实现的1~2 ×10- 5 moloL - 1。产生阻遏蛋白的基因是trpR ,该基因距trp o2peron基因簇很远。它结合于trp 操纵基因特异序列,阻止转录起始。但阻遏蛋白的DNA结合活性受Trp调控, Trp起着一个效应分子的作用, Trp与之结合的动力学常数为。在有高浓度Trp存在时,阻遏蛋白- 色氨酸复合物形成一个同源二聚体,并且与色氨酸操纵子紧密结合,因此可以阻止转录。阻遏蛋白- 色氨酸复合物与基因特异位点结合的能力很强,动力学常数为2 ×10- 10mol·L - 1 ,因此细胞内阻遏蛋白数量仅有20~30分子已可充分发挥作用。当Trp 水平低时,阻遏蛋白以一种非活性形式存在,不能结合DNA。在这样的条件下, trp操纵子被RNA聚合酶转录,同时Trp 生物合成途径被激活。
弱化作用
trp操纵子转录终止的调控是通过弱化作用(衰减作用)( attenuation)实现的。为什么trp操纵子需要这种额外的调控机制?可能的原因在于trp操纵子的阻遏作用实质上是一种弱调控机制,即使有阻遏物存在的情况下,trp操纵子仍能发生大量的转录事件。事实上,在阻遏调控系统和衰减调控系统的共同作用下,可以在700倍左右范围内对trp操纵子的转录活性进行调控。这十分经济有效,因为色氨酸的合成需要消耗大量地能量。
在大肠杆菌trp operon,前导区的碱基序列包括4个分别以1、2、3和4表示的片段,能以两种不同的方式进行碱基配对, 1 - 2和3 -4配对,或2 - 3配对, 3 - 4配对区正好位于终止密码子的识别区。前导序列有相邻的两个色氨酸密码子,当培养基中Trp 浓度很低时,负载有Trp 的tR2NATrp也就少,这样翻译通过两个相邻色氨酸密码子的速度就会很慢,当4区被转录完成时,核糖体滞留1区,这时的前导区结构是2 - 3配对,不形成3 - 4配对的终止结构,所以转录可继续进行。反之,核糖体可顺利通过两个相邻的色氨酸密码子,在4区被转录之前,核糖体就到达2区,这样使2 - 3不能配对, 3 - 4 区可以配对形成终止子结构, 转录停止。枯草杆菌的弱化作用机制另有特点。因其色氨酸操纵子结构的特殊性,转录起始的调节似乎不如转录终止的调节更具重要性。枯草杆菌色氨酸操纵子表达主要受到色氨酸激活RNA结合蛋白( Trp -activated RNA - binding p rotein, TRAP)的调节。该蛋白与色氨酸结合被激活后,可与trpE上游转录产物结合, 导致转录终止。当色氨酸浓度较低时,TRAP失活,转录可以继续,结构基因得以表达。另外枯草杆菌对未负荷色氨酸的tRNATrp也很敏感,后者大量堆积, 会诱导合成抗TRAP 蛋白( anti -PRAP, AT) 。AT与Trp激活的PRAP结合,可以取消其转录终止活性。trpG表达也受PRAP调控,活化的TRAP与和trpG相重叠的S - D 序列结合,阻碍核糖体的结合,抑制trpG转录。
反馈抑制
由于基因表达必然消耗一定的能源和前体物,相对于阻遏和弱化作用,反馈抑制作用更为经济和高效。终产物Trp对催化分支途径几步反应的酶具有反馈抑制作用,其50%抑制浓度分别为:邻氨基苯甲酸合酶, 0. 0015 mmol·L - 1 ;邻氨基苯甲酸磷酸核糖转移酶, 0. 15 mmol·L - 1 ; 色氨酸合成酶, 7. 7mmol·L - 1 。对于普通野生菌株,邻氨基苯甲酸合酶对Trp合成起到关键调控作用,常被称为瓶颈酶;但对高产Trp工程菌而言,上述任何一种酶的反馈抑制都会直接影响Trp产量。研究发现酶蛋白某些特殊位点突变可以导致对反馈抑制作用敏感性显著下降,如邻氨基苯甲酸合酶38位的丝氨酸被精氨酸取代,抗反馈抑制能力显著提高,当环境中Trp浓度为10 mmol·L - 1时酶活性不受影响,而相同条件下野生型酶活性不到1%。邻氨基苯甲酸磷酸核糖转移酶162位缬氨酸被谷氨酸取代,抗反馈抑制能力也有显著提高,当环境中含有0. 83 mmol·L - 1色氨酸或0. 32 mmol·L - 1 5 - 甲基- 色氨酸时,酶活性分别为野生菌的3. 6倍和2. 4倍。陈小芳等报道一株谷氨酸棒杆菌邻氨基苯甲酸合酶基因7个碱基突变导致6个氨基酸残基改变,抗反馈抑制能力显著增强,环境中Trp 浓度达到15 mmol·L - 1时,邻氨基苯甲酸合酶活性几乎没有变化。
遗传改造
由于色氨酸操纵子的调控作用,自然界不可能存在高产Trp菌株,为了获得高产Trp菌株,就必须对色氨酸操纵子进行改造,解除其调节作用。早期的研究策略主要依靠传统诱变方法,经过长期努力,获得了一些有价值的研究结果,如获得了TrpR - 菌株,通过缺失某些片断解除了弱化作用,得到了一些抗反馈抑制的酶。许多Trp生产菌株都是通过随机的诱变技术筛选得到的,如王健等通过硫酸二乙酯诱变, Trp 类似物筛选等方法从谷氨酸棒杆菌中培育出一株trp 高产菌株,摇瓶发酵64 h,产trp达到7. 28 g·L - 1。
传统诱变的方法尽管有效,但其缺陷点也是显而易见的,如工作量大,效率低,突变株的菌体生长、对环境的耐受性以及遗传稳定性等都比野生型菌株差等。基因工程技术的建立和发展对色氨酸操纵子改造提供了新的技术平台。1979年Tribe等人采用DNA重组技术对大肠杆菌进行改造,扩增trp 操纵子,发酵12 h,产酸1 g·L - 1 ,产酸量尽管不是很高,但是其意义却十分重大,由此开创了基因工程技术在Trp生物合成应用的先河。随后,Aiba等将带有色氨酸操纵子的质粒引入大肠杆菌,发酵27 h,并补充邻氨基苯甲酸,得到trp 6. 2 g·L - 1 。Ikeda等通过构建稳定质粒,扩增分支途径限速酶并改造中心代谢途径, 获得产Trp 达58 mg ·L - 1 的菌株。除了扩增表达操纵子基因,对其进行理性设计和改造也开始引起关注。已知酶分子某些特殊位点突变可以导致对反馈抑制作用敏感性下降,因此可以考虑利用基因工程技术对色氨酸操纵子结构基因进行理性改造降低其对反馈抑制的敏感性,但是目前尚缺乏成功的范例,主要原因在于现有酶分子反馈抑制结构与功能关系资料不足,不能满足需要。[1][3][4]


-

-

-

如何看待汽车价格 如何看待汽车降价后又涨价的现象呢?
2025-11-02 00:07:17 查看详情 -
如何看待年后汽车价格趋势 如何看待汽车降价后又涨价的现象呢?
2025-11-02 00:07:17 查看详情 -
如何确定汽车价格上涨了 如何理解汽车降价后的涨价现象?
2025-11-02 00:07:17 查看详情 -
发动机漏油 发动机表面有漏油现象,严重吗?
2025-11-02 00:07:17 查看详情 -
汽车同步器坏了有什么现象
2025-11-02 00:07:17 查看详情 -

-

-


 求购
求购