- 巨椎龙

巨椎龙
简介
恐龙名称: 巨椎龙/大脊椎龙(大陆)
拉丁文名: Mssospondylus
恐龙体长: 5米
恐龙体重: 不详
恐龙食物: 草食
生存年代: 侏罗纪早期
生存地点: 北美和非洲南部
辨认要决: 拇指长有长而弯曲的爪
恐龙种类: ?
恐龙脖子:长
基本内容
巨椎龙是最早在陆地上出现的以植物为食的恐龙之一。它的头很小,脖子和尾巴却很长。依靠两条后腿直立起来时, 它能够到大树顶上的嫩芽和树叶。巨椎龙的牙齿很小,可以咬碎树叶,但咀嚼功能却不强。当这种恐龙的化石被初步发现的时候,在它的肋骨笼内找到了一些小卵石。科学家们估计这是大椎龙吞下帮助它们在胃中消化食物的。卵石可以将树叶捣碎成浓厚、粘稠的汁液,以便恐龙能够吸收对身体有用的营养。巨椎龙的拇指特别大,上面长有长而弯曲的爪,主要是为了防御。在二、三指的配合下,大拇指还具有抓握功能。另外两个指则又小又弱。
它能够到大树顶上的嫩芽和树叶。巨椎龙的牙齿很小,可以咬碎树叶,但咀嚼功能却不强。当这种恐龙的化石被初步发现的时候,在它的肋骨笼内找到了一些小卵石。科学家们估计这是大椎龙吞下帮助它们在胃中消化食物的。卵石可以将树叶捣碎成浓厚、粘稠的汁液,以便恐龙能够吸收对身体有用的营养。巨椎龙的拇指特别大,上面长有长而弯曲的爪,主要是为了防御。在二、三指的配合下,大拇指还具有抓握功能。另外两个指则又小又弱。
叙述
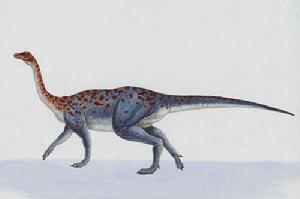 大椎龙是种中等大小的原蜥脚类恐龙,身长约4米,体重接近135公斤,少数的研究则估计大椎龙的身长可达6米。大椎龙长久以来被认为是四足动物,但2007年的一份对于前肢生理构造的研究,则认为从它们的动作范围,排除了惯常四足步伐的可能。这个研究也认为大椎龙手部转动的幅度有限,排除了以指关节着地或其他形式的行走方式。
大椎龙是种中等大小的原蜥脚类恐龙,身长约4米,体重接近135公斤,少数的研究则估计大椎龙的身长可达6米。大椎龙长久以来被认为是四足动物,但2007年的一份对于前肢生理构造的研究,则认为从它们的动作范围,排除了惯常四足步伐的可能。这个研究也认为大椎龙手部转动的幅度有限,排除了以指关节着地或其他形式的行走方式。
在其他方面,大椎龙是种典型的原蜥脚类恐龙。它们的身体修长,颈部长,具有大约9节长颈椎、13节背椎、3节荐椎、以及至少40节尾椎。耻骨朝前,如同大部分的蜥臀目恐龙。与同为原蜥脚类的板龙相比,大椎龙的身体较为轻型。一个近年的发现,显示大椎龙具有发展良好的锁骨,并连接成类似叉骨的型态,由此可知它们的肩胛骨固定不动,更可知这些锁骨不像那些没有真正叉骨的恐龙一样地缺乏功能。这个发现也指出鸟类的叉骨是从锁骨演化而来的。
如同板龙,大椎龙的每个脚掌都有5根脚趾,拇指有大型指爪,可用来协助进食,或抵御掠食者。前掌的第四与第五指小型,使前掌看起来不对称。该2007年的研究指出大椎龙的双手手掌面朝内;而化石出土时,腕部从未保持在关节连接的状态。
发现
大椎龙的模式种是刀背大椎龙(M. carinatus),是由理查·欧文所建立。属名衍化自古希腊文,masson/μάσσων意为“巨大的”,而sphondylos/σφονδυλος意为“脊椎”;而种名意为“剃刀般的背部”[20]。在2009年,保罗·巴雷特(Paul Barrett)将一个发现于南非上艾略特组的部分头颅骨,命名为新种M. kaalae。上艾略特组也发现过一些刀背大椎龙的化石,显示这两个种生存于相同年代。这两个种的头颅骨形态、特征不同,例如M. kaalae的前上颌骨比例较长[21]。
大椎龙过去曾有过许多其他种,但大部分不被承认是有效种,包括:
布朗氏大椎龙(M. browni):由哈利·丝莱(H.G. Seeley)在1895年命名。
哈里斯氏大椎龙(M. harriesi):由罗伯特·布鲁姆(Robert Broom)在1911年命名。
希氏大椎龙(M. hislopi):由理查德·莱德克(Richard Lydekker)在1890年命名。
休尼氏大椎龙(M. huenei): 由库柏(M. R. Crooper)在1981年命名。
拉氏大椎龙(M. rawesi):由莱德克在1890年命名。
施氏大椎龙(M. schwarzi): 由席尼·贺顿(Sydney H. Haughton)在1924年命名。
布朗氏大椎龙、哈里斯氏大椎龙、以及施氏大椎龙,都发现于南非角省的上艾略特组。三者都只有破碎的化石,大部分近年的研究将它们视为疑名。布朗氏大椎龙的化石有2节颈椎、2节背椎、3节尾椎、以及混杂的后肢碎片。哈里斯氏大椎龙的化石只有前肢。施氏大椎龙的化石是一个不完整后肢与荐骨。希氏大椎龙与拉氏大椎龙的化石都发现于印度。希氏大椎龙的化石为脊椎,发现于安得拉邦的马勒尔组,年代为晚三叠纪。拉氏大椎龙的化石是两颗牙齿,发现于马哈拉施特拉邦的Takli组,年代为晚白垩纪。在最近的看法里,希氏大椎龙暂时被视为一种无法确定的蜥脚形亚目动物。拉氏大椎龙可能是种兽脚类恐龙,或者并非恐龙。库柏则提出休尼氏大椎龙就是许氏禄丰龙,因此他认为禄丰龙与大椎龙是同种动物。但这个看法并不被接受。
疑名
有数种恐龙常被认为是大椎龙的异名,包括:Aristosaurus、Dromicosaurus、Gryponyx、Hortalotarsus、Leptospondylus、以及Pachyspondylus,这些恐龙的有效性不大,也被视为疑名。Hortalotarsus是由哈利·丝莱在1894年建立,化石包含部份腿骨。第二年,理察·欧文将一些脊椎命名为Leptospondylus与Pachyspondylus。这些化石都在第二次世界大战期间遭到摧毁。在1920年,E.C.N. van Hoepen根据一个接近完整的骨骸,建立了Aristosaurus。他另外将一个部份骨骸命名为Dromicosaurus。在1924年,席尼·贺顿命名了Gryponyx,化石为臀部骨头。以上化石都发现于南非的赫塘阶到锡内穆阶,约跟大椎龙同一时期。根据国际动物命名法规,这些名称是大椎龙的次同物异名。大椎龙比这些名称还早出现在科学文献中,因此大椎龙具有优先权。
分类
巨椎龙是早期食草恐龙。全长4-5米,头小颈长,外形比同时期的板龙要小巧得多。一般四脚着地,也能仅用后腿站 立起来采食。前肢上的"手"很大,拇指上长着大而弯曲的爪,这种结构的手可能用来捡取树叶。人们从这种恐龙的化石中发现,它除了吃树的枝叶外,还时常吞食些鹅卵石,很可能它的牙齿不足以嚼碎食物,只能把这些石头放在胃里充当碾磨器。这种办法传递给了后来的一些大型食草恐龙,甚至今天的鸟类。
立起来采食。前肢上的"手"很大,拇指上长着大而弯曲的爪,这种结构的手可能用来捡取树叶。人们从这种恐龙的化石中发现,它除了吃树的枝叶外,还时常吞食些鹅卵石,很可能它的牙齿不足以嚼碎食物,只能把这些石头放在胃里充当碾磨器。这种办法传递给了后来的一些大型食草恐龙,甚至今天的鸟类。
大椎龙属于原蜥脚下目,原蜥脚下目是群早期蜥臀目恐龙,生存于三叠纪与侏罗纪,并在侏罗纪末期灭亡。原蜥脚下目的其他著名属包含:板龙、云南龙、与里奥哈龙。基础蜥脚形亚目的系统发生学仍在争论中,根据亲缘分支分类法,一个天然演化支应包含它们的共同祖先与其共同祖先的所有后代,所以许多过去被认为是典型原蜥脚类的物种,因为无法与原有物种构成一天然演化支,近年被排除在原蜥脚下目之外。但哪些物种构成原蜥脚类为一个单系群,仍不确定。在2003年,亚当·耶茨(Adam Yates)与詹姆斯·基钦(James Kitching)公布了一个演化支,包含:里奥哈龙、板龙、科罗拉多斯龙、大椎龙、以及禄丰龙。在2004年,彼得·加尔东(Peter M.Galton)与保罗·阿普彻奇(Paul Upchurch)则将原蜥脚下目列为单系群,包含:砂龙、近蜥龙、爱珍多龙、卡米洛特龙、科罗拉多斯龙、优肢龙、金山龙、莱森龙、禄丰龙、大椎龙、黑丘龙、鼠龙、板龙、里奥哈龙、吕勒龙、农神龙、鞍龙、槽齿龙、易门龙、以及云南龙。在2005年,杰佛瑞·威尔森(Jeffrey A. Wilson)提出大椎龙、金山龙、板龙、与禄丰龙,形成一个天然演化支,可能还有贝里肯龙、雷前龙等蜥脚类恐龙。在2007年,Matthew F. Bonnan与耶茨提出卡米洛特龙、贝里肯龙、黑山龙可能属于蜥脚下目。同样在2007年,耶茨将雷前龙、黑丘龙、贝里肯龙归类为基础蜥脚类恐龙,而且认为原蜥脚下目是板龙科的异名,而没有使用这分类。但耶茨并没有排除少部份原蜥脚类构成一个单系群的可能性,这些成员包含板龙、里奥哈龙、大椎龙、以及它们的最近亲。
古生态学
如同所有恐龙,大椎龙的许多生物学层面仍然未知,例如:行为、外表颜色、生理机能。但是近年的研究提出了关于生长模式、食性、步态、繁衍、以及呼吸等方面的假设。
一个2007年的研究提出大椎龙会将短前肢用在抵抗掠食动物、物种内打斗、或是协助进食,但它们的前肢太短,无法碰到嘴部。科学家们假设大椎龙会使用拇指指爪来打斗,或是折下树上的食物。


-

380TSI劲擎智联版四驱车型上市 成都车展:雪铁龙C
2025-09-22 16:39:58 查看详情 -
1万元首付 云南雪铁龙C2最低首付1.88万元
2025-09-22 16:39:58 查看详情 -
成都日产天籁冠军版综合优惠5.5万 贵阳雪铁龙C3
2025-09-22 16:39:58 查看详情 -
成都雪铁龙C6最高优惠6万元 新凯美瑞全系现金优惠6千
2025-09-22 16:39:58 查看详情 -
郑州长安CS75优惠高达1.6万 北京雪铁龙C5现车
2025-09-22 16:39:58 查看详情 -
MAX优惠3000元 北京雪铁龙C5现车
2025-09-22 16:39:58 查看详情 -
贷款购车低首付低利率 成都雪铁龙C2按揭首付2万元
2025-09-22 16:39:58 查看详情 -
云南雪铁龙C2最低首付1.88万元 0首付换购天籁开回家
2025-09-22 16:39:58 查看详情 -
雪铁龙凡尔赛C5 新款东风雪铁龙天逸C5
2025-09-22 16:39:58 查看详情 -
一汽 一汽丰田新款亚洲龙申报图
2025-09-22 16:39:58 查看详情

 求购
求购