- 线虫动物门

线虫动物门
形态特征
线虫动物,体细长圆柱状,故又称圆虫。体表被一层较厚的角质膜,蛋白质成分,为上皮所分泌,一般分为皮层、中层和基层3层,有保护作用。角质膜下为合胞体的上皮,即上皮的细胞界限不清,具多核。上皮向内突起成纵脊,两侧的为侧线,背面和腹面的为背线和腹线。线虫体内有一广阔的空腔,称为初生体腔或假体腔(pseudocoelom),是胚胎时期的囊腔发展而来,整个腔没有完全被中胚层所覆盖,即只有体壁中胚层,且不具体腔膜。腔内充满体腔液,致使虫体鼓胀饱满,身体难以任意伸缩。肌肉层位上皮下,为一层纵肌,不发达,故身体只能弯曲作波浪状蠕动。线虫具有完整的消化道,即有口和肛门,与腔肠动物和扁形动物相比,是个很大的进步。消化道管状,分前肠、中肠和后肠。前、后肠来源于外胚层,无消化作用,中肠为内胚层来源,是消化吸收的主要部分。
 线虫动物门(3)
线虫动物门(3)
神经系统
 线虫动物门(3)线虫在咽的周围有一环状的脑,环的两侧膨大成神经节,由脑环向前后各分出六条神经,前端的神经分布到唇、乳突及化感器等。向后的六条神经中,一条为背神经,一条为腹神经,二对侧神经,两对侧神经离开脑环后很快合并成一对,最后的这四条神经分别位于相应的纵行上皮索内,其中腹神经最发达,由腹神经发出分支到肠及肛门。腹神经索中包括运动神经纤维及感觉神经纤维,背神经索中主要为运动神经纤维,侧神经索中主要为感觉神经纤维。另外在脑环周围,神经细胞集中,形成神经节状。
线虫动物门(3)线虫在咽的周围有一环状的脑,环的两侧膨大成神经节,由脑环向前后各分出六条神经,前端的神经分布到唇、乳突及化感器等。向后的六条神经中,一条为背神经,一条为腹神经,二对侧神经,两对侧神经离开脑环后很快合并成一对,最后的这四条神经分别位于相应的纵行上皮索内,其中腹神经最发达,由腹神经发出分支到肠及肛门。腹神经索中包括运动神经纤维及感觉神经纤维,背神经索中主要为运动神经纤维,侧神经索中主要为感觉神经纤维。另外在脑环周围,神经细胞集中,形成神经节状。
 线虫动物门(3)线虫的感觉器官主要分布在头部及尾部两端。头部包括唇、乳突、感觉毛,统称为头感器;还有一个特殊的化感器(amphid)。唇及乳突是头部的角质突起,有脑环发出的神经进行支配。感觉毛在头部较发达是一种触觉感受器,实际上是一种改变了的纤毛,有的种感觉毛周围还有腺细胞围绕。化感器是线虫特有的一种感觉器,位于身体前端一侧,它是体表的一个内陷物,呈囊状、管状、螺旋状等各种形态,成为线虫分目的依据之一,其内端为盲端,外端开口,它是一种化学感受器,也常有腺细胞伴随。电子显微镜的研究已经证明化感器的感觉突实际上也是改变了的纤毛,过去一直认为线虫类不存在任何纤毛,而化感器是改变了的纤毛,这种特征便将线虫和其他有纤毛的动物在进化上联系起来。在水生种类特别是海产种类化感器发达,而陆生及寄生种类则退化了。水生种类在咽的两侧还有一对眼点,是视觉器官其中色素细胞分散或排列成杯状。
线虫动物门(3)线虫的感觉器官主要分布在头部及尾部两端。头部包括唇、乳突、感觉毛,统称为头感器;还有一个特殊的化感器(amphid)。唇及乳突是头部的角质突起,有脑环发出的神经进行支配。感觉毛在头部较发达是一种触觉感受器,实际上是一种改变了的纤毛,有的种感觉毛周围还有腺细胞围绕。化感器是线虫特有的一种感觉器,位于身体前端一侧,它是体表的一个内陷物,呈囊状、管状、螺旋状等各种形态,成为线虫分目的依据之一,其内端为盲端,外端开口,它是一种化学感受器,也常有腺细胞伴随。电子显微镜的研究已经证明化感器的感觉突实际上也是改变了的纤毛,过去一直认为线虫类不存在任何纤毛,而化感器是改变了的纤毛,这种特征便将线虫和其他有纤毛的动物在进化上联系起来。在水生种类特别是海产种类化感器发达,而陆生及寄生种类则退化了。水生种类在咽的两侧还有一对眼点,是视觉器官其中色素细胞分散或排列成杯状。
线虫的尾端也有一对单细胞腺体称尾感器(phasmid),分别开口在尾端两侧,它也是一种腺状感受器,这种感受器在寄生的种类发达。雄性个体尾端交配器周围也有感觉乳突及感觉毛。
体壁运动
 线虫动物门(3)线虫的体壁亦由角质层、上皮细胞及肌肉组成。线虫的角质层结构较复杂,可分为三层,其最外层为一层鞣化蛋白质,表现出环纹;中层为均质的,内层为胶原蛋白构成的支持柱层。寄生的蛔虫类等,其内层由于纤维排列的方向不同,又可分为三层,因此使角质层表现出一定的弹性。角质层是上皮细胞的分泌物,它限制了身体的生长,因此线虫在生长过程中经过数次到多次的脱皮现象。在脱皮前,角质层中有用的物质被虫体吸收,然后上皮细胞重新分泌新的表皮,使旧表皮与上皮细胞分离,并最后完全脱落,使虫体得以生长。研究指明,线虫的脱皮是由神经环上的神经细胞分泌出激素(hormone),激素促使排泄细胞分泌脱皮液,脱皮液可溶解旧表皮而造成脱皮,脱皮现象仅出现在幼虫期,成虫仅能增加角质层的厚度而不再脱皮。角质层内为上皮细胞,上皮细胞或界线清楚或为合胞体。在身体背、腹中线及两侧、上皮细胞向内凸出形成4条纵行的上皮索。上皮细胞核仅局限在索中,并排列成行。这4条上皮索在体表明显可见,分别称为背线、腹线及侧线。上皮之内为肌肉层,线虫缺乏环肌,只有纵肌,分布在上皮索之间,肌肉为斜纹肌。肌细胞的基部为可收缩的肌纤维,端部为不能收缩的细胞体部,它的功能可能是贮存糖元,核位于细胞体部。细胞体部的原生质延伸形成线状,分别连接到背索与腹索内的神经上,在那里接受神经支配。其他动物是由神经发出分支分布到肌肉上进行支配,而不是像线虫由肌肉延伸到神经处,去接受支配。体壁之内为假体腔,其中充满体腔液,体腔液内没有游离的细胞,但有体腔细胞固着在肠壁及体壁上,体腔液除了担任输送营养物及代谢物之外,还有抗衡肌肉收缩所产生的压力,起着骨骼的作用。线虫的运动是由纵肌的收缩及角质层的弹性改变而共同完成。当背纵肌收缩时,腹面角质层中的纤维拉长,当背纵肌松弛时,腹面角质层中的纤维恢复,因此表现出身体背腹方向的蛇行运动。一些种类体表具刺、环等,可做短距离的爬行或游泳运动。
线虫动物门(3)线虫的体壁亦由角质层、上皮细胞及肌肉组成。线虫的角质层结构较复杂,可分为三层,其最外层为一层鞣化蛋白质,表现出环纹;中层为均质的,内层为胶原蛋白构成的支持柱层。寄生的蛔虫类等,其内层由于纤维排列的方向不同,又可分为三层,因此使角质层表现出一定的弹性。角质层是上皮细胞的分泌物,它限制了身体的生长,因此线虫在生长过程中经过数次到多次的脱皮现象。在脱皮前,角质层中有用的物质被虫体吸收,然后上皮细胞重新分泌新的表皮,使旧表皮与上皮细胞分离,并最后完全脱落,使虫体得以生长。研究指明,线虫的脱皮是由神经环上的神经细胞分泌出激素(hormone),激素促使排泄细胞分泌脱皮液,脱皮液可溶解旧表皮而造成脱皮,脱皮现象仅出现在幼虫期,成虫仅能增加角质层的厚度而不再脱皮。角质层内为上皮细胞,上皮细胞或界线清楚或为合胞体。在身体背、腹中线及两侧、上皮细胞向内凸出形成4条纵行的上皮索。上皮细胞核仅局限在索中,并排列成行。这4条上皮索在体表明显可见,分别称为背线、腹线及侧线。上皮之内为肌肉层,线虫缺乏环肌,只有纵肌,分布在上皮索之间,肌肉为斜纹肌。肌细胞的基部为可收缩的肌纤维,端部为不能收缩的细胞体部,它的功能可能是贮存糖元,核位于细胞体部。细胞体部的原生质延伸形成线状,分别连接到背索与腹索内的神经上,在那里接受神经支配。其他动物是由神经发出分支分布到肌肉上进行支配,而不是像线虫由肌肉延伸到神经处,去接受支配。体壁之内为假体腔,其中充满体腔液,体腔液内没有游离的细胞,但有体腔细胞固着在肠壁及体壁上,体腔液除了担任输送营养物及代谢物之外,还有抗衡肌肉收缩所产生的压力,起着骨骼的作用。线虫的运动是由纵肌的收缩及角质层的弹性改变而共同完成。当背纵肌收缩时,腹面角质层中的纤维拉长,当背纵肌松弛时,腹面角质层中的纤维恢复,因此表现出身体背腹方向的蛇行运动。一些种类体表具刺、环等,可做短距离的爬行或游泳运动。
取食消化
 线虫动物门(3)线虫的食性很广泛,许多自由生活的线虫是肉食性的,以小形的动物为食,也有许多种为植食性的,以藻类及植物根部细胞及其内含物为食,还有的种类是细菌及沉积物的取食者,是以溶解的动、植物尸体或有机颗粒为食,它们构成了数量最大、分布最广的细菌及真菌的取食者,在生物链中起着重要作用。线虫的消化系统结构简单,为一直形管,前端口后为一管状或囊状的口囊(buccal capsule),口囊内壁角质层加厚,形成不同形状或不同数目的嵴、板、齿等结构,用以切割食物。特别在肉食性的种类较发达。有的种类在口囊中形成一中空或实心的刺,用以穿刺食物或抽吸食物汁液。口囊之后为咽,咽常形成一个或几个咽球,由于肌肉细胞的加厚,咽腔在断面上呈三放形,三放中的一放总是指向腹中线,构成线虫咽的一个特征。咽的周围有成对的咽腺,可分泌消化液,咽腺可开口在咽前端。咽由于很厚的肌肉层,具有泵的作用,可由口抽吸食物进入咽及肠。咽后紧接为中肠,是由单层上皮细胞组成,中肠的两端均有瓣膜,以阻止肠内食物逆流。中肠后为短的直肠,最后以肛门开口在近末端的腹中线上。线虫的咽腺及中肠的腺细胞产生消化酶,在中肠内进行食物的消化,并在肠壁细胞内完成细胞内的消化。线虫类或通过体表进行气体交换或行低氧呼吸,它没有专门的呼吸及循环结构。
线虫动物门(3)线虫的食性很广泛,许多自由生活的线虫是肉食性的,以小形的动物为食,也有许多种为植食性的,以藻类及植物根部细胞及其内含物为食,还有的种类是细菌及沉积物的取食者,是以溶解的动、植物尸体或有机颗粒为食,它们构成了数量最大、分布最广的细菌及真菌的取食者,在生物链中起着重要作用。线虫的消化系统结构简单,为一直形管,前端口后为一管状或囊状的口囊(buccal capsule),口囊内壁角质层加厚,形成不同形状或不同数目的嵴、板、齿等结构,用以切割食物。特别在肉食性的种类较发达。有的种类在口囊中形成一中空或实心的刺,用以穿刺食物或抽吸食物汁液。口囊之后为咽,咽常形成一个或几个咽球,由于肌肉细胞的加厚,咽腔在断面上呈三放形,三放中的一放总是指向腹中线,构成线虫咽的一个特征。咽的周围有成对的咽腺,可分泌消化液,咽腺可开口在咽前端。咽由于很厚的肌肉层,具有泵的作用,可由口抽吸食物进入咽及肠。咽后紧接为中肠,是由单层上皮细胞组成,中肠的两端均有瓣膜,以阻止肠内食物逆流。中肠后为短的直肠,最后以肛门开口在近末端的腹中线上。线虫的咽腺及中肠的腺细胞产生消化酶,在中肠内进行食物的消化,并在肠壁细胞内完成细胞内的消化。线虫类或通过体表进行气体交换或行低氧呼吸,它没有专门的呼吸及循环结构。
排泄调节
 线虫动物门(4)线虫的排泄器官属于原肾型,而结构不典型,是由腺型细胞或由细胞形成的管完成排泄功能。原肾型特点是由外胚层内陷形成;只在体表有开口,体内没有开口。但这些动物的代谢废物更多的是从体表排出,因此原肾排泄系统的主要功能是保持体液的稳定,保持动物体内的水盐平衡(这与原生动物伸缩泡的作用类似)。原始的种类只有1~2个大型的腺细胞进行排泄及水分调节,这种腺细胞具有长颈,位于咽的周围,也称肾细胞(renette cell),联合开口在神经环附近的腹中线上,如小杆线虫。另一些种,腺型肾细胞延伸成管型排泄器,多呈H型管,两个侧管在前端以一横管相连分别位于侧上皮索内,最后经过共同的小管以排泄孔开口在前端腹中线上,例如驼形线虫(Camallanus)。蛔虫(Ascaris)的排泄管也呈H形,只是横行管成网状,侧管前端不发达,而且是由一个腺细胞特化形成。线虫的腺细胞及排泄管中没有鞭毛或纤毛,不呈焰细胞状。线虫的代谢产物主要是氨,可以通过体壁及消化道而排出。线虫的排泄器官对维持其体腔液的压力是十分重要的,水可以通过口及体壁进入体内,过多的水分可通过排泄器官排出。实验证明:海产的种类可以在氯化钠的低渗液中进行调节,而不能在高渗液中调节;淡水及陆生的种类相反,维持体腔液低渗于周围环境;陆生线虫还可以在高度干燥条件下,像轮虫一样通过失水,降低代谢速率,处于隐生状态,来渡过干燥可长达数年之久。
线虫动物门(4)线虫的排泄器官属于原肾型,而结构不典型,是由腺型细胞或由细胞形成的管完成排泄功能。原肾型特点是由外胚层内陷形成;只在体表有开口,体内没有开口。但这些动物的代谢废物更多的是从体表排出,因此原肾排泄系统的主要功能是保持体液的稳定,保持动物体内的水盐平衡(这与原生动物伸缩泡的作用类似)。原始的种类只有1~2个大型的腺细胞进行排泄及水分调节,这种腺细胞具有长颈,位于咽的周围,也称肾细胞(renette cell),联合开口在神经环附近的腹中线上,如小杆线虫。另一些种,腺型肾细胞延伸成管型排泄器,多呈H型管,两个侧管在前端以一横管相连分别位于侧上皮索内,最后经过共同的小管以排泄孔开口在前端腹中线上,例如驼形线虫(Camallanus)。蛔虫(Ascaris)的排泄管也呈H形,只是横行管成网状,侧管前端不发达,而且是由一个腺细胞特化形成。线虫的腺细胞及排泄管中没有鞭毛或纤毛,不呈焰细胞状。线虫的代谢产物主要是氨,可以通过体壁及消化道而排出。线虫的排泄器官对维持其体腔液的压力是十分重要的,水可以通过口及体壁进入体内,过多的水分可通过排泄器官排出。实验证明:海产的种类可以在氯化钠的低渗液中进行调节,而不能在高渗液中调节;淡水及陆生的种类相反,维持体腔液低渗于周围环境;陆生线虫还可以在高度干燥条件下,像轮虫一样通过失水,降低代谢速率,处于隐生状态,来渡过干燥可长达数年之久。
生殖发育
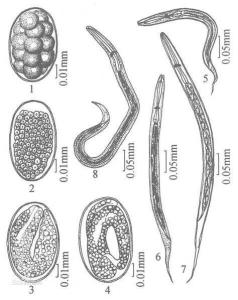
 线虫动物门线虫主要为雌雄异体及异形,雄性个体较小,后端弯曲成钩状。生殖腺均为管状,生殖细胞是由生殖腺中一个大的末端细胞(terminal cell)发生,在其通过生殖腺的过程中,生长成熟。少数线虫生殖细胞来自整个生殖腺的管壁细胞。雄性生殖系统通常具有一个精巢,少数种是两个。后端连接输精管及其膨大的贮精囊,向后为肌肉发达的射精管(ejaculatory duct),开口到直肠或泄殖腔。射精管周围有数目不等的前列腺(prostatic gland)。大多数线虫雄性泄殖腔向外伸出两个囊,每个囊中有一角质的交合刺(spicule),有肌肉牵引可自由的由泄殖孔伸出体外交配时用以撑开阴门。交合刺的长短、形态因种而异,也是分类标准之一。还有的线虫交合刺的背壁有角质小骨片,愈合成副刺(gubernaculum),以控制交合刺的运动。线虫的精子具鞭毛或不具鞭毛而呈囊状、球状等。雌性个体通常有两个卵巢,少数个体仅一个卵巢,卵巢管长短不等,相对排列或平行排列,卵巢后端为输卵管、子宫。子宫的上端为受精囊,用以贮存交配后的精子,两个子宫后端联合,经肌肉质阴道,以雌性生殖孔开孔在身体近中部腹中线上。线虫需交配受精,交配时雄虫以尾端对准雌性生殖孔,再以交合刺撑开阴门,由交合囊及射精管肌肉的收缩送精子入雌体,精子经鞭毛运动或变形运动到达子宫上端,在此与卵融合,卵受精后形成厚的受精膜,变硬后形成卵的内壳,卵沿子宫下行时,子宫的分泌物形成卵的外壳,卵壳外层的形状常作为寄生线虫分类的依据。卵常在子宫内时已开始发育。极少数的线虫是雌雄同体的,如小杆线虫目的一些种,并行自体受精。也有极少数陆生线虫可行孤雌生殖。自由生活的线虫产卵量较少,海产种类一般仅产数十粒卵,陆生种产卵较多,可达数百粒。寄生种类产卵量极大,每日可产卵数千到数十万粒。线虫的卵是决定型卵,分裂球排列不对称,许多种细胞分裂在胚胎期已经完成,除了生殖系统之外,其他的器官细胞数已固定,孵化后幼虫的生长是通过细胞体积的增加而实现。幼虫期一般脱皮4次,前两次脱皮常在卵壳中进行,成年后不再脱皮。
线虫动物门线虫主要为雌雄异体及异形,雄性个体较小,后端弯曲成钩状。生殖腺均为管状,生殖细胞是由生殖腺中一个大的末端细胞(terminal cell)发生,在其通过生殖腺的过程中,生长成熟。少数线虫生殖细胞来自整个生殖腺的管壁细胞。雄性生殖系统通常具有一个精巢,少数种是两个。后端连接输精管及其膨大的贮精囊,向后为肌肉发达的射精管(ejaculatory duct),开口到直肠或泄殖腔。射精管周围有数目不等的前列腺(prostatic gland)。大多数线虫雄性泄殖腔向外伸出两个囊,每个囊中有一角质的交合刺(spicule),有肌肉牵引可自由的由泄殖孔伸出体外交配时用以撑开阴门。交合刺的长短、形态因种而异,也是分类标准之一。还有的线虫交合刺的背壁有角质小骨片,愈合成副刺(gubernaculum),以控制交合刺的运动。线虫的精子具鞭毛或不具鞭毛而呈囊状、球状等。雌性个体通常有两个卵巢,少数个体仅一个卵巢,卵巢管长短不等,相对排列或平行排列,卵巢后端为输卵管、子宫。子宫的上端为受精囊,用以贮存交配后的精子,两个子宫后端联合,经肌肉质阴道,以雌性生殖孔开孔在身体近中部腹中线上。线虫需交配受精,交配时雄虫以尾端对准雌性生殖孔,再以交合刺撑开阴门,由交合囊及射精管肌肉的收缩送精子入雌体,精子经鞭毛运动或变形运动到达子宫上端,在此与卵融合,卵受精后形成厚的受精膜,变硬后形成卵的内壳,卵沿子宫下行时,子宫的分泌物形成卵的外壳,卵壳外层的形状常作为寄生线虫分类的依据。卵常在子宫内时已开始发育。极少数的线虫是雌雄同体的,如小杆线虫目的一些种,并行自体受精。也有极少数陆生线虫可行孤雌生殖。自由生活的线虫产卵量较少,海产种类一般仅产数十粒卵,陆生种产卵较多,可达数百粒。寄生种类产卵量极大,每日可产卵数千到数十万粒。线虫的卵是决定型卵,分裂球排列不对称,许多种细胞分裂在胚胎期已经完成,除了生殖系统之外,其他的器官细胞数已固定,孵化后幼虫的生长是通过细胞体积的增加而实现。幼虫期一般脱皮4次,前两次脱皮常在卵壳中进行,成年后不再脱皮。
生活习性
线虫的生活环境和生活史是复杂、多样的,概括来说,可以分为自由生活的、腐生的和寄生的三大类。营自由生活的,水生的种类一般都营底栖生活,海产的最多,由海滩到极深的海底都有,有时数目众多,成为海底多细胞动物数量最大的一类,淡水生活的种类存在于各种各样的淡水环境中,甚至在急流中也可以见到,常用其尾腺分泌的粘液固着在水中物体上;在温泉水温高达37℃的环境里,有时也可以见到某些种类。陆生的种类常生活在土壤、树根、落叶下。有的线虫甚至在水生和陆生中均可以生活,属于一种两栖的类型。显然线虫起源于海洋,经两栖生活,再进入到陆地,从区系分类的角度看,海产和淡水产的种类几乎完全不同,因此淡水产的种类很可能是由海产的种类先进入陆地,然后再侵入淡水水域的。营寄生生活的种类,可寄生于动物或植物的体内,有的成虫营寄生生活,而幼虫营自由生活;或有的幼虫营寄生生活,而成虫营自由生活;或为幼虫、成虫都营寄生生活,形形色色,不一而足。至于营腐生生活的,则是一群生活在动、植物尸体内,或以死亡后的动、植物躯体为食的类群。可以认为,腐生生活是从自由生活过渡到寄生生活的中间阶段,如杆形虫属Rhabditis的许多种类,以动、植物的尸体为食,而某些杆形虫能行厌氧呼吸,因此它们能通过动物的消化道而对己无害,在肠道内不增殖,但也不被消化不致死亡。
繁殖饲养
自由生活的线虫通过体表进行呼吸,寄生种类为厌氧呼吸。排泄器官在自由生活种类,由1~2大细胞称原肾细胞(renettecell)完成,称腺型排泄器官。寄生线虫具“H”型排泄管,是原肾细胞特化形成,称管型排泄器官。生殖器官,雄性有精巢、输精管、储精囊、射精管各1个,开口于泄殖腔(cloaca),以肛门开口于体外。雌性有卵巢、输卵管、子宫各1对。由阴道以雌性生殖孔开口于体表。生殖由雌雄交配,卵在体内受精。自由生活种类产卵量少,寄生种类产卵量巨大,有的一条雌虫每天产卵数10万个。神经为神经环、神经节和神经干组成。神经环位咽部,连于神经节,由神经环向前后伸出数条神经干。感官有刚毛、乳突、化感器(amphid)等。线虫头端有刚毛,口具唇,唇上有乳突;头端有侧器(lateralorgan),尾端有乳突或尾感器(phasmid)。
种类介绍
 线虫动物门线虫的分类,尚有分歧,分目标准也各不相同,根据Maggenti(1981)的分类将线虫分为2个纲、20目。
线虫动物门线虫的分类,尚有分歧,分目标准也各不相同,根据Maggenti(1981)的分类将线虫分为2个纲、20目。
无尾感器纲(Aphasmida)或有腺纲(Adenophorea)身体尾端无尾感器,有尾腺,排泄器官腺状,由单细胞或多细胞腺体组成,海水、淡水、土壤及动植物体内均有分布,海产线虫仅限于本纲。
尾感器纲(Phasmida)或胞管肾纲(Secernentea)身体尾端具一对尾感器,无尾腺,化感器不发达,排泄器官为胞管状,位于身体两侧上皮索内。咽腺通常3个,绝大多数为陆生,偶然在淡水中发现,无海产种,分为3个亚纲。


-

-

人类与动物心理学论稿
2025-09-22 01:43:26 查看详情 -

长城汽车发布生命体征监测技术 或仍将以动物命名
2025-09-22 01:43:26 查看详情 -
-
-

-

-

-
西汉虎耳动物格斗铜贮贝器
2025-09-22 01:43:26 查看详情 -


 求购
求购