- 古菌

古菌
形态
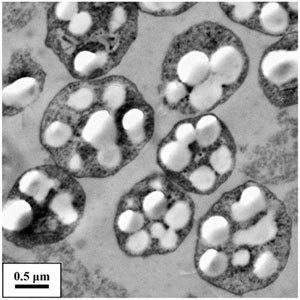 古菌的细胞形态有球形、杆状、螺旋形、耳垂形、盘状、不 规则形状、多形态,有的很薄、扁平,有的有精确的方角和垂直的边构成直角几何形态,有的以单个细胞存在,有的呈丝状体或 团聚体。其直径大小一般在0.1~15μm, 丝状体长度有200μm。
古菌的细胞形态有球形、杆状、螺旋形、耳垂形、盘状、不 规则形状、多形态,有的很薄、扁平,有的有精确的方角和垂直的边构成直角几何形态,有的以单个细胞存在,有的呈丝状体或 团聚体。其直径大小一般在0.1~15μm, 丝状体长度有200μm。
细胞结构
古菌的细胞结构与细菌不同,如古菌的细胞外膜就与细菌不同。
大多数古菌的细胞壁不含 二氨基庚二酸(D- 氨基酸)和 胞壁酸,不受 溶菌酶和内酰胺抗生素如青霉素的作用。革兰氏阳性古菌的细胞壁含有各种复杂的多聚体,如产甲烷菌的细胞壁含 假肽聚糖,甲烷八叠 球菌和盐球菌不含假肽聚糖,而含复杂聚多糖。革兰氏阴性古菌没有外膜,含蛋白质或 糖蛋白亚基的表层,其厚度在20~40nm。 甲烷叶菌属、盐 杆菌属和极端嗜热的 硫化叶菌属、热 变形菌属和 热网菌属的细胞壁有 糖蛋白; 甲烷球菌属、 甲烷微菌属、 产甲烷菌属和极端嗜热的脱硫球菌属有蛋白质壁,蛋白质呈酸性。
古菌的细胞膜所含脂质与细菌的很不同,细菌的脂类是 甘油脂肪酸酯,而古菌的脂质是非皂化性甘油二醚的磷脂和 糖脂的衍生物。古菌的细胞膜有两种:双层膜和单层膜。
代谢
古菌在代谢过程中,有许多特殊的辅酶。古菌因有5个类群,所以,它们的代谢呈多样性。古菌中有异养型、 自养型和不完全光合作用3种类型。
古菌多数为严格厌氧、兼性厌氧,还有专性好氧。
 古菌
古菌
繁殖方式
古菌的繁殖方式有二分裂、芽殖。其繁殖速度较慢, 进化速度也比细菌慢。
生活习性
大多数古菌生活在极端环境,如盐分高的湖泊水中,极热、极酸和绝对厌氧的环境,有的在极冷的环境生存。
分类
按照古菌的生活习性和生理特性,古菌可分为三大类型:产甲烷菌、嗜热嗜酸菌、极端嗜盐菌。
从2001年起着手出版的《伯杰系统细菌学手册》第二版中,将原核生物分为 古菌域和 细菌域。 古菌域分类为泉 古生菌门和广古生菌门。
泉古生菌门
大多数泉古生菌门极端嗜热、嗜酸,代谢硫。硫在 厌氧呼吸中作为 电子受体和无机营养的电子源。它们多生长在 含硫地热水或土壤中(如 美国的 黄石国家公园的富硫 温泉)。
广古生菌门
1.产甲烷菌
2.极端嗜盐菌
3.热原体
4.古生 硫酸盐还原菌
5.嗜热嗜酸菌
发现
“古细菌”这个概念是1977年由卡尔·沃斯和George Fox提出的,原因是它们在16SrRNA的系统发生树上和其它原核生物的区别。这两组原核生物起初被定为古细菌(Archaebacteria)和 真细菌(Eubacteria)两个界或 亚界。Woese认为它们是两支根本不同的生物,于是重新命名其为古菌(Archaea)和细菌(Bacteria),这两支和真核生物(Eukarya)一起构成了生物的三域系统。
如果将地球约46亿年的年龄比作一年,那么古菌早在3月20日就出现了,而人类诞生不过是12月31日的事。它们多生长于极端环境,如热泉、高压的海底 火山口、盐湖等。
人们对 古菌的兴趣并非始于1970年代。事实上,在我们这个星球上,古菌代表着 生命的极限,确定了 生物圈的范围。例如,一种叫做热网菌(Pyrodictium)的古菌能够在高达113℃的温度下生长。这是迄今为止发现的最高生物生长温度。近年来,利用 分子生物学方法,人们发现,古菌还广泛分布于各种自然环境中, 土壤、 海水、 沼泽地中均生活着古菌。
目前,可在 实验室培养的古菌主要包括三大类: 产甲烷菌、 极端嗜热菌和 极端嗜盐菌。产甲烷菌生活于富含有机质且严格无氧的环境中,如 沼泽地、水稻田、 反刍动物的 反刍胃等,参与地球上的 碳素循环,负责甲烷的 生物合成;极端嗜盐菌生活于 盐湖、 盐田及盐腌制品表面,它能够在盐饱和环境中生长,而当盐浓度低于10%时则不能生长;极端嗜热菌通常分布于 含硫或硫化物的 陆相或水相地质热点,如含 硫的 热泉、 泥潭、海底热溢口等,绝大多数极端嗜热菌严格 厌氧,在获得 能量时完成硫的转化。
尽管生活习性大相径庭,古菌的各个类群却有共同的、有别于其他 生物的细胞学及生物化学特征。例如,古菌 细胞膜含由分枝碳氢链与D型磷酸甘油,以醚键相连接而成的脂类,而细菌及真核生物细胞膜则含由不分枝脂肪酸与L型磷酸甘油,以酯键相连接而成的脂类。细菌 细胞壁的主要成分是 肽聚糖,而古菌细胞壁不含肽聚糖。
有趣的是,虽然与 细菌相似,古菌 染色体 DNA呈闭合环状, 基因也组织成操纵子(操纵子为原核生物 基因表达和调控的基本结构 单位, 生物活性相关的基因常以操纵子的结构形式协调基因表达的开启和关闭),但在DNA复制、转录、翻译等方面,古菌却具有明显的真核特征:采用非甲 酰化甲硫 氨酰tRNA作为 起始tRNA, 启动子、 转录因子、 DNA聚合酶、 RNA聚合酶等均与 真核生物的相似。
比较 生物化学的研究结果表明, 古菌与细菌有着本质的区别,这种区别与两者表现在 系统发育学方面亲缘关系的疏远是一致的。
两种学说
地球上究竟有几种生命形式?当 亚里士多德建立生物学时,他用二分法则将生物分为动物和植物。显微镜的诞生使人们发现了肉眼看不见的细菌。细菌在 细胞结构上与动植物的最根本差别是,动植物细胞内有细胞核, 遗传物质DNA主要储存于此,而细菌则没有细胞核,DNA游离于 细胞质中。由于动物与植物的差别小于它们与细菌的差别,沙东(E. Chatton)于1937年提出了生物界新的二分法则,即生物分为含 细胞核的 真核生物和不含细胞核的 原核生物。动植物属于 真核生物,而细菌属于 原核生物。
1859年 达尔文发表《 物种起源》以后,生物学家便开始建立基于进化关系而非表型相似性的 分类系统,即所谓系统发育分类系统。可是,由于缺乏 化石记录,这种 分类方法长期未能有效运用于原核生物的分类。1970年代,随着分子生物学的发展,伍斯终于在这方面获得了意义重大的突破。
在漫长的进化过程中,每种生物细胞中的 信息分子( 核酸和蛋白质)的序列均不断发生着突变。许多信息分子序列变化的产生在时间上是随机的, 进化速率相对恒定,即具有时钟特性。因此,物种间的亲缘关系可以用它们共有的某个具有时钟特性的基因或其产物(如 蛋白质)在序列上的差别来定量描述。这些基因或其产物便成了记录生物进化历程的分子记时器(chronometer)。显然,这种记录 生物系统发育历程的分子记时器应该广泛分布于所有生物之中。基于这一考虑,伍斯选择了一种名为 小亚基 核糖体核酸(SSU rRNA)的分子,作为分子记时器。这种分子是细胞内 蛋白质合成机器—— 核糖体的一个组成部分,而蛋白质合成又是几乎所有生物 生命活动的一个重要方面。因此,把SSU rRNA分子作为 分子记时器是合适的。
在比较了来自不同原核及真核生物的SSU rRNA序列的相似性后,伍斯发现原先被认为是细菌的甲烷 球菌代表着一种既不同于真核生物,也不同于细菌的 生命形式。考虑到甲烷 球菌的生活环境可能与生命诞生时地球上的自然环境相似,伍斯将这类生物称为 古细菌。据此,伍斯于1977年提出,生物可分为三大类群,即真核生物、 真细菌和古细菌。基于SSU rRNA分析结果的泛系统发育(进化)树随后诞生了。
进一步的研究表明,进化树上的第一次分叉产生了 真细菌的一支和古细菌/真核生物的一支,古细菌和真核生物的分叉发生在后。换句话说,古细菌比 真细菌更接近真核生物。
据此,1990年伍斯提出了三域分类学说:生物分为真核生物、 真细菌和古细菌三域,域被定义为高于界的分类单位。为突出古细菌与 真细菌的区别,伍斯将古细菌更名为古菌。 真细菌改称细菌。 三域学说使古菌获得了与真核生物和细菌等同的分类学地位。
自诞生之日起,伍斯的 三域学说便遭到部分人,特别是 微生物学领域外的人反对。反对者坚持认为:原核与真核的区分是生物界最根本的、具有进化意义的分类法则;与具有丰富 多样性表型的真核生物相比,古菌与 细菌的差异远没有大到需要改变二分法则的程度。但在 詹氏甲烷球菌基因组序列测定完成前的近20年中,采用多种分子记时器进行的系统发育学研究一再证明,古菌是一种独特的 生命形式。
三域学说
尽管对古菌已有了上述认识,当人们第一次面对詹氏甲烷球菌的全基因组序列时,还是大吃了一惊。詹氏甲烷球菌共有1738个基因,其中人们从未见过的基因竟占了56%!相比之下,在这之前完成测序的流感嗜血菌(Haemophilus influenzae)和生殖道枝 原体(Mycoplasma genitalium)基因组中未知基因仅占20%左右。于是人们终于在基因组水平上认识到,古菌是一种崭新的 生命形式。
更有趣的是,詹氏甲烷球菌基因组中占总数44%的那些功能 或多或少已经知道的基因似乎勾勒出了古菌与另两类生物之间的进化关系:古菌在产能、 细胞分裂、代谢等方面与细菌相近,而在转录、翻译和复制方面则与真核生物类似。换言之,一个生活在 大洋底部热溢口处的、习性古怪的微生物,在遗传信息传递方面竟有着与人(而不是与人的消化道中细菌)相似的基因!在赞叹生命奇妙的同时,许多人开始欢呼 三域学说的最终确立。美国《科学》周刊在把詹氏甲烷球菌基因组序列测定工作列为1996年度重大科学突破之一时宣称,这一成果使围绕 三域学说的争论差不多可以结束了。
新挑战
就在古菌的悬念似乎行将消失时,接踵而来的新发现却使人们重新陷入困惑之中。各类完整的 微生物基因组序列一个接一个出现在人们轻点鼠标便可查阅的数据库中,在已发表的18种生物基因组序列中,古菌的占了4个。采用更灵敏的方法对这些基因组(包括詹氏甲烷球菌基因组)进行分析,得到了令人吃惊的结果:詹氏甲烷球菌基因组中只有30%(不是先前估计的半数以上)的基因编码目前未知的功能,这与细菌基因组相近。古菌的神秘性和独特性因此减少了许多。
对 三域学说更为不利的是,在詹氏甲烷球菌的那些可以推测功能的 基因产物(蛋白质)中,44%具有细菌蛋白特征,只有13%的像真核生物的蛋白质。在另一个古菌,嗜热碱 甲烷杆菌(Methanobacterium thermoaotutrophicum)的基因组中也有类似情况。因此,从基因组比较的数字上看,古菌与细菌间的差异远小于古菌与真核生物间的差异,不足以说服 三域学说的反对者。
更令人难以理解的是,利用同一生物中不同基因对该物种进行系统发育学定位常常会得到不同的结果。最近,一种能在接近沸点温度下生长的细菌(Aquifex aeolicus)的基因组序列测定完成。对该菌的几个基因进行的系统发育学研究表明:如果用参与 细胞分裂调控的蛋白质FtsY作为分子记时器,该菌与伍斯进化树上位于细菌分枝的一个 土壤细菌—— 枯草芽孢杆菌相近;如果以一种参与色氨酸合成的酶为准,该菌应属于古菌;而当比较该菌和其他生物的合成 胞苷三磷酸(DNA的基本结构单位之一)的酶时,竟发现古菌不再形成独立的一群。看来不同的基因似乎在诉说不同的进化故事。那么,古菌还能是独特的、统一的 生命形式吗?
属于真核生物的啤酒酵母基因组序列测定完成后, 三域学说遇到了更大危机。酵母细胞核基因中,与细菌基因有亲缘关系的比与古菌有亲缘关系的多一倍。有人还对在三种 生命形式中都存在的34个 蛋白质家族进行了分析,发现其中17个家族来源于细菌,只有8个显示出古菌与真核生物的亲缘关系。
如果伍斯进化树正确、古菌与真核生物在进化历程中的分歧晚于两者与细菌的分歧的话,那么怎样才能解释上面这些结果呢?
根据细胞进化研究中流行的内共生假说, 真核细胞 细胞器( 线粒体、 叶绿体)的产生源于细菌与原真核生物在进化早期建立的内共生关系。在这种关系中, 真核细胞提供稳定的微环境, 内共生体(细菌)则提供 能量,久而久之,内共生体演变为 细胞器。真核生物细胞核中一部分源于细菌的基因可能来自 线粒体,这些为数不多的基因通常编码重新运回线粒体的蛋白质分子。可是,现在发现许多源于细菌的 核基因编码那些在 细胞质、而不是线粒体中起作用的蛋白质。那么,这些基因从何而来呢?显然,内共生假说已不足以挽救伍斯进化树。
不过,伍斯进化树也不会轻易倒下,支撑它的假说依然很多。最近,有人提出了新版的“ 基因水平转移”假说。根据这个假说,基因组的杂合组成是进化过程中不同谱系间发生 基因转移造成的。一种生物可以采用包括吞食等方式获得另一种、亲缘关系也许很远的生物的基因。伍斯推测, 始祖生物在演化形成细菌、古菌和真核生物三大谱系前,生活于可以相互交换基因的“公社”中,来自这个“史前公社”的生物可能获得了不同的基因遗产。这一切使得进化树难以枝杈分明。不过,伍斯相信,基于SSU rRNA的进化树在总体上是正确的,三种 生命形式是存在的。
争论
三年前詹氏甲烷球菌基因组序列的发表,似乎预示着一场延续了20多年的、关于地球上到底有几种生命形式的争论的终结。古菌似乎被认定为生命的第三种形式。如今,仅仅过了三年,即使最乐观的人都无法预料伍斯进化树的命运。这场仍在继续的争论中,尽管古菌的分类地位遭到质疑,但古菌这一生命形式的独特性依然得到不同程度的肯定。
目前,古菌研究正在世界范围内升温,这不仅因为古菌中蕴藏着远多于另两类生物的、未知的生物学过程和功能,以及有助于阐明 生物进化规律的线索,而且因为古菌有着不可估量的生物技术开发前景。古菌已经一次又一次让人们吃惊,可以肯定,在未来的岁月中,这群独特的生物将继续向人们展示生命的无穷奥秘。




 求购
求购